
Potentialité post activité PAP
Primo rédacteur: Lucas Bourgeois 2023
Définition de la Post Activation Potentiation :
La Post-Activation-Potentiation (PAP) est définie comme un état neuromusculaire amélioré, observé après l'exécution d'un exercice de haute intensité1. La PAP se définie par une augmentation de la force de contraction du muscle qui suit une contraction volontaire maximale ou quasi maximale. Le but étant d’entrainer une plus grande production de force si le muscle a été stimulé, activé au préalable.
Les mécanismes physiologiques de la PAP :
D’un point de vue plus physiologique, l’augmentation de la force de contraction musculaire serait liée à une augmentation de la phosphorylation de la chaîne légère de myosine rendant l’actine – myosine plus sensible au calcium CA2+ libéré par le réticulum sarcoplasmique au moment de la contraction musculaire2,3. En conséquence, la force de contraction de chaque fibre musculaire s'en voit augmentée. On suppose également que les adaptations de l'activité réflexe dans la moelle épinière, après l'exécution d'un exercice de haute intensité, contribuent à la réponse PAP4,5,6,7. Ainsi, il a été proposé que deux mécanismes physiologiques soient responsables de la PAP. L’un est la phosphorylation des chaines légères régulatrice de myosine et l’autre est une augmentation du recrutement des unités motrices d’ordre supérieur.
Phosphorylation des chaines légères régulatrice de myosine (RLC) :
Une molécule de myosine est un hexamère composé de deux chaînes lourdes (figure 1)8. Les extrémités aminées de chaque chaîne lourde, classées comme tête de myosine, contiennent deux RLC, et chaque RLC possède un site de liaison spécifique pour l'incorporation d'une molécule de phosphate. La phosphorylation des RLC est catalysée par l'enzyme kinase de la chaîne légère de myosine, qui est activée lorsque les molécules de Ca2+, libérées du réticulum sarcoplasmique pendant la contraction musculaire, se lient à la calmoduline, une protéine régulatrice du calcium.
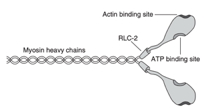
Figure 1 : Une molécule de myosine. Chaque molécule de myosine est composée de deux chaînes lourdes de myosine. La chaîne légère régulatrice (RLC)-2 représente une paire de RLC positionnées au niveau du cou d'une tête de myosine. Chaque RLC peut incorporer une molécule de phosphate, ce qui modifie la structure de la tête de la myosine. Chaque tête de myosine comporte un site de liaison à l'actine et à l'adénosine triphosphate (ATP)8.
Les chercheurs suggèrent que la phosphorylation du RLC potentialise les contractions subséquentes en modifiant la structure de la tête de la myosine et en l'éloignant de son filament épais. La phosphorylation du RLC a son plus grand effet à des concentrations relativement faibles de Ca2+. Relativement peu d'études ont tenté de mesurer la réponse PAP dans le muscle squelettique humain mais Stuart et al. (1988)9 ont enregistré une teneur en phosphate significativement élevée du RLC dans le muscle vaste latéral (p <0,01), et une potentialisation significative de la tension de commutation des extenseurs du genou, après une contraction volontaire maximale isométrique de 10 secondes (MVC ; p <0,05). Il y avait également une corrélation positive mais non significative entre l'étendue de la potentialisation de la contraction et la quantité de phosphate incorporée dans les unités RLC individuelles, et entre la potentialisation et le pourcentage de fibres musculaires de type II (p >0,05)9.
Recrutement des unités motrices d'ordre supérieur :
Les recherches ont montré qu'une contraction isométrique provoquée par la stimulation de fibres neurales afférentes spécifiques, qui activent à leur tour les a motoneurones adjacents via une augmentation neuronale, augmente la transmission des potentiels d'excitation à travers les jonctions synaptiques au niveau de la moelle épinière. Cet état d'accommodation peut durer plusieurs minutes après la contraction isométrique et il en résulte une augmentation des potentiels post-synaptiques pour le même potentiel pré-synaptique lors d'une activité ultérieure. Les chercheurs pensent qu'une contraction musculaire induite diminue la défaillance du transmetteur au cours de l'activité suivante, par l'intermédiaire d'une ou d'une combinaison de plusieurs réponses possibles. Il s'agit notamment d'une augmentation de la quantité de neurotransmetteur libéré, d'une augmentation de l'efficacité du neurotransmetteur ou d'une réduction de la défaillance du point de ramification axonal le long des fibres neuronales afférentes8.
Modification de la courbe force-vitesse :
Dans la plupart des activités sportives explosives, la puissance joue un rôle majeur et la puissance mécanique peut être définie comme la vitesse à laquelle la force est développée sur une plage de mouvement, dans une période de temps spécifique, ou comme la force multipliée par la vitesse. L'augmentation du niveau de force à une vitesse donnée permet d’augmenter la puissance mécanique. Bien que la force maximale ou la vitesse maximale, situées aux extrémités de la courbe force-vitesse ne veut pas être modifiés par la PAP, la PAP peut tout de même augmenter la vitesse de développement de la Force même à des fréquences de stimulation très élevées où la force n’est pas augmentée par la PAP. En augmentant le taux de développement de la force et donc l'accélération atteinte avec des charges entre zéro et la force maximale, la PAP augmente la vitesse atteinte avec ces charges. Ainsi, la PAP déplacerait la relation charge (force)-vitesse vers le haut et vers la droite, la rendant moins concave, sans modifier les extrémités, comme le montre la figure 2.
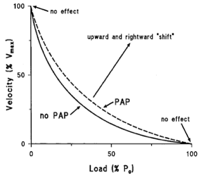
Figure 2. La relation force vitesse modifiée par la PAP3.
Ici, P0 représente la Force maximale et Vmax la vitesse maximale de raccourcissement.
PAP ou PAPE ?
Les adaptations décrites ci-dessus sont souvent désignés comme un effet de « post activation potentiation » (PAP). Nous avons vu que l’activité de conditionnement stimulait un afflux de calcium sarcoplasmique (Ca2+) dans le myoplasme qui active la kinase de la chaîne de myosine, qui phosphoryle les chaînes légères et favorise le cycle des ponts actine-myosine en raison d’une plus grande sensibilité au Ca2+, entrainant des améliorations de force. Physiologiquement, cette amélioration dure moins de 30 secondes10, tandis que les améliorations de production de force se produisent souvent entre 6 et 10 minutes selon la littérature. Ainsi, lorsque des modifications de la force volontaire sont observées, le terme amélioration de la performance post-activation (PAPE) a récemment été proposé dans plusieurs études10,11,12. Les mécanismes alternatifs proposés pour la PAPE incluent des augmentations de la température musculaire, de l’activation musculaire, ou des altérations du schéma moteur. Ces deux appellations sont proposées dans le tableau 1. Pour l’ensemble du document nous garderons l’appellation PAP.
Tableau 1. Présentation des mécanismes de PAP et PAPE10 ,11.
|
|
PAP |
PAPE |
|
Post-Activation Potentiation |
Post-Activation Performance Enhancement |
|
|
Effet de la performance |
Amélioration des propriétés de la contraction musculaire |
Amélioration de la production de force dynamique volontaire |
|
Déclenchement de la potentiation |
Effets immédiats et transitoires, démarre dans les secondes qui suivent l’activité de conditionnement (environ 30s) |
Effets retardés et durables, démarre dans les minutes qui suivent l’activité de conditionnement et est opérationnelle entre 6-10 minutes |
|
Mécanismes de Potentiation |
Phosphorylation de la chaine légère régulatrice de myosine (MRLC)
|
1- L’augmentation de la température musculaire est associée à une augmentation de la vitesse de développement de la force et de la vitesse de raccourcissement dans les muscles/fibres à contraction rapide et lente. 2- l'augmentation du flux sanguin et de l'eau intracellulaire peut améliorer la production de force musculaire, effet plus important dans les fibres de type II. 3- L'augmentation du niveau de la commande neurale volontaire des contractions de conditionnement peut améliorer l'activation musculaire, qui augmente le taux maximal de développement de la force et la force musculaire maximale. |
|
Inhibition |
Fatigue et interférence des schémas moteurs |
Fatigue et interférence des schémas moteurs |
L’influence de la période de repos entre l’AC et la performance suivante sur la PAP :
Le temps de récupération entre l’échauffement PAP et l’activité de puissance suivante est un facteur important qui influence la réponse de potentiation. Il faut choisir un temps suffisamment long mais pas trop pour permettre aux effets de la fatigue de se dissiper sans perdre la potentialisation ou l'effet PAP2,13.
Si la fatigue est trop importante, par exemple si le sprint avait lieu immédiatement après le mouvement de PAP, alors la PAP ne pourrait pas avoir d'effets optimaux1. Et de même, si trop de temps s'écoule, la fatigue diminue, mais les effets de la PAP aussi.
Plusieurs études ont examiné l'impact de différents temps de récupération sur la performance des activités de puissance. Ces études ont trouvé des impacts positifs sur la performance des activités de puissance avec des périodes de récupération allant de 0 à 20 minutes14,15,16,17.
Une période de repos appropriée après un échauffement PAP peut également dépendre des activités concernées. Par exemple, certains chercheurs18 ont constaté une amélioration des temps de sprint sur 100 m après que les participants aient bénéficié d'un temps de récupération de 9 minutes. Des études sur 30m, 40m ont utilisé un temps de récupération de 4 à 5 minutes19,20. Ces études ont utilisé un squat lourd comme mouvement PAP dans l'échauffement. Bien que plusieurs chercheurs aient examiné le concept d'échauffement PAP, la plupart des études ont porté sur des exercices tels que les squats et les sauts verticaux, plutôt que sur des épreuves de compétition comme le sprint. Les études qui ont démontré un effet significatif de l'échauffement PAP sur la performance du sprint ont utilisé une période de repos de 4 à 10 minutes.
L’équilibre entre la PAP et la fatigue et son effet sur les contractions explosives ultérieures ont été observé et ont rapporté que la fatigue semble plus dominante dans les premiers stades de la récupération et, par conséquent, la performance de l'activité volontaire subséquente est diminuée ou inchangée. Cependant, la fatigue s'estompe plus rapidement que la PAP, et une potentialisation des performances peut être réalisée à un moment donné de la période de récupération. La figure 3 illustre la relation PAP-fatigue.
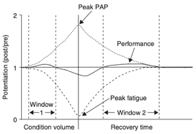
Figure 3 : Modèle de la relation entre la potentialisation post-activation (PAP) et la fatigue à la suite d'un protocole de contraction de pré-conditionnement. Lorsque le volume de la condition est faible, la PAP est plus dominante que la fatigue, et une potentialisation de la performance explosive ultérieure (post/pre) peut être réalisée immédiatement (window 1). Au fur et à mesure que le volume de la condition augmente, la fatigue devient dominante, affectant négativement les performances ultérieures. Après la condition, la fatigue se dissipe plus rapidement que la PAP, et une potentialisation de la performance explosive ultérieure peut être réalisée à un moment donné pendant la période de récupération (window 2)8.
Si les contractions isométriques et dynamiques peuvent induire des réponses différentes à la fatigue, on peut supposer qu'elles peuvent également avoir des effets différents sur les mécanismes de la PAP : les CMV isométriques peuvent induire une fatigue centrale, mais une PAP périphérique, tandis que les CMV dynamiques peuvent induire la réponse inverse.
Zisi et al (2022)21, dans leur étude sur les effets de l’échauffement en résistance avec une charge correspondant à une réduction de 50 % de la vitesse maximale théorique de l'individu (entre 57 et 73 % de la masse corporelle), ont utilisé une récupération de 5 minutes après l'échauffement et avant le premier sprint de 30m.
Dans l’étude de Winwood et al (2016)22, les principales conclusions de cette étude sont que la charge de 75 % sur le charriot a entraîné de légères améliorations des temps de sprint du groupe de 5, 10 et 15 m après 8 et 12 minutes de récupération. La récupération de 8 minutes entre l'activité de conditionnement et la performance ultérieure utilisée dans cette étude est suffisante pour minimiser la fatigue et obtenir l'effet PAP. Ce résultat est cohérent avec l’étude de Jarvis et al (2017)23 qui a démontré des temps de sprint plus rapides sur 15 m après des sprints résistants avec une charge de 50% de BM et 8 min de récupération.
Un exemple d’application sur le terrain avec la charge de travail et le temps de récupération favorisant un effet PAP en sprint avec résistance est présenté en annexes.
Relation entre PAP et fibres musculaires
Une amélioration des performances grâce à un échauffement PAP dépend aussi de la composition des fibres musculaires. Plus les athlètes ont un pourcentage élevé de fibres musculaires à contraction rapide ou de type II, plus ils présentent des réponses plus importantes à la PAP tandis que les athlètes ayant un pourcentage plus élevé de fibre lente, de type I sont moins sujet à répondre à cet effet PAP6. Les fibres II produisent une plus grande quantité de phosphorylation de la chaîne légère de la myosine face à une activité intense, ce qui rend l’actine-myosine plus sensible au calcium lors des contractions musculaires suivantes.
Les individus les plus susceptibles de bénéficier de la PAP sont ceux qui ont une plus grande force musculaire, un pourcentage plus élevé de fibres de type 2 (bien que la fatigue puisse également être plus importante chez ces individus), et un rapport puissance-force plus faible, en effet, les sujets les moins capables de convertir efficacement leur force en puissance sont plus susceptibles de bénéficier de la PAP que ceux qui le peuvent.
Bibliographie:
1Robbins, D. W. (2005). Postactivation potentiation and its practical applicability : A brief review. Journal of Strength and Conditioning Research, 19(2), 453‑458. https://doi.org/10.1519/R-14653.1
2Hodgson, M., Docherty, D., & Robbins, D. (2005). Post-activation potentiation : Underlying physiology and implications for motor performance. Sports Medicine (Auckland, N.Z.), 35(7), 585‑595. https://doi.org/10.2165/00007256-200535070-00004
3Sale, D. G. (2002). Postactivation potentiation : Role in human performance. Exercise and Sport Sciences Reviews, 30(3), 138‑143. https://doi.org/10.1097/00003677-200207000-00008
4Chiu, L. Z. F., Fry, A. C., Weiss, L. W., Schilling, B. K., Brown, L. E., & Smith, S. L. (2003). Postactivation potentiation response in athletic and recreationally trained individuals. Journal of Strength and Conditioning Research, 17(4), 671‑677. https://doi.org/10.1519/1533-4287(2003)017<0671:ppriaa>2.0.co;2
5Guellich, A., & Schmidtbleicher, D. (1996). MVC-induced short-term potentiation of explosiv force. New Studies in Athletics, 11, 67‑84.
6Hamada, T., Sale, D. G., MacDougall, J. D., & Tarnopolsky, M. A. (2000). Postactivation potentiation, fiber type, and twitch contraction time in human knee extensor muscles. JournalofAppliedPhysiology,88(6),2131‑2137.https://doi.org/10.1152/jappl.2000.88.6.2131
7Scott, S., & Docherty, D. (2004). Acute Effects of Heavy Preloading on Vertical and Horizontal Jump Performance. Journal of strength and conditioning research / National Strength & Conditioning Association, 18, 201‑205. https://doi.org/10.1519/R-13123.1
8Tillin, N., & Bishop, D. J. (2009). Factors Modulating Post-Activation Potentiation and its Effect on Performance of Subsequent Explosive Activities. Sports medicine (Auckland, N.Z.), 39, 147‑166. https://doi.org/10.2165/00007256-200939020-00004
9Stuart, D. S., Lingley, M. D., Grange, R. W., & Houston, M. E. (1988). Myosin light chain phosphorylation and contractile performance of human skeletal muscle. Canadian Journal of Physiology and Pharmacology, 66(1), 49‑54. https://doi.org/10.1139/y88-009
10Blazevich, A. J., & Babault, N. (2019). Post-activation Potentiation Versus Post-activation Performance Enhancement in Humans : Historical Perspective, Underlying Mechanisms, and Current Issues. Frontiers in Physiology, 10, 1359. https://doi.org/10.3389/fphys.2019.01359
11Cormier, P., Freitas, T. T., Loturco, I., Turner, A., Virgile, A., Haff, G. G., Blazevich, A. J., Agar-Newman, D., Henneberry, M., Baker, D. G., McGuigan, M., Alcaraz, P. E., & Bishop, C. (2022). Within Session Exercise Sequencing During Programming for Complex Training : Historical Perspectives, Terminology, and Training Considerations. Sports Medicine, 52(10), 2371‑2389. https://doi.org/10.1007/s40279-022-01715-x
12Prieske, O., Behrens, M., Chaabene, H., Granacher, U., & Maffiuletti, N. A. (2020). Time to Differentiate Postactivation “Potentiation” from “Performance Enhancement” in the Strength and Conditioning Community. Sports Medicine, 50(9), 1559‑1565. https://doi.org/10.1007/s40279-020-01300-0
13Weber, K. R., Brown, L. E., Coburn, J. W., & Zinder, S. M. (2008). Acute effects of heavy-load squats on consecutive squat jump performance. Journal of Strength and Conditioning Research, 22(3), 726‑730. https://doi.org/10.1519/JSC.0b013e3181660899
14Bevan, H., Cunningham, D., Tooley, E., Owen, N., Cook, C., & Kilduff, L. (2010). Influence of Postactivation Potentiation on Sprinting Performance in Professional Rugby Players. Journal of strength and conditioning research / National Strength & Conditioning Association, 24, 701‑705. https://doi.org/10.1519/JSC.0b013e3181c7b68a
15Comyns, T. M., Harrison, A. J., Hennessy, L. K., & Jensen, R. L. (2006). The optimal complex training rest interval for athletes from anaerobic sports. Journal of Strength and Conditioning Research, 20(3), 471‑476. https://doi.org/10.1519/18445.1
16Docherty, D., Robbins, D., & Hodgson, M. (2004). Complex Training Revisited : A Review of its Current Status as a Viable Training Approach. Strength & Conditioning Journal, 26. https://doi.org/10.1519/1533-4295(2004)026<0052:CTRARO>2.0.CO;2
17Kilduff, L., Bevan, H., Kingsley, M., Owen, N., Bennett, M., Bunce, P., Hore, A., Maw, J., & Cunningham, D. (2007). Postactivation Potentiation in Professional Rugby Players : Optimal Recovery. Journal of strength and conditioning research / National Strength & Conditioning Association, 21, 1134‑1138. https://doi.org/10.1519/R-20996.1
18Linder, E. E., Prins, J. H., Murata, N. M., Derenne, C., Morgan, C. F., & Solomon, J. R. (2010). Effects of preload 4 repetition maximum on 100-m sprint times in collegiate women. Journal of Strength and Conditioning Research, 24(5), 1184‑1190. https://doi.org/10.1519/JSC.0b013e3181d75806
19Chatzopoulos, D., Michailidis, C., Giannakos, A., Alexiou, K., Patikas, D., Antonopoulos, C., & Kotzamanidis, C. (2007). Postactivation Potentiation Effects After Heavy Resistance Exercise on Running Speed. Journal of strength and conditioning research / National Strength & Conditioning Association, 21, 1278‑1281. https://doi.org/10.1519/R-21276.1
20McBride, J. M., Nimphius, S., & Erickson, T. M. (2005). The acute effects of heavy-load squats and loaded countermovement jumps on sprint performance. Journal of Strength and Conditioning Research, 19(4), 893‑897. https://doi.org/10.1519/R-16304.1
21Zisi, M., Stavridis, I., Agilara, G.-O., Economou, T., & Paradisis, G. (2022). The Acute Effects of Heavy Sled Towing on Acceleration Performance and Sprint Mechanical and Kinematic Characteristics. Sports (Basel, Switzerland), 10(5), 77. https://doi.org/10.3390/sports10050077
22Winwood, P., Posthumus, L., Cronin, J., & Keogh, J. (2016). The Acute Potentiating Effects of Heavy Sled Pulls on Sprint Performance. The Journal of Strength and Conditioning Research, In Press. https://doi.org/10.1519/JSC.0000000000001227
23Jarvis, P., Turner, A., Chavda, S., & Bishop, C. (2017). The Acute Effects of Heavy Sled Towing on Subsequent Sprint Acceleration Performance. Journal of Trainology, 6. https://doi.org/10.17338/trainology.6.1_18
Annexes :
Tableau présentant la charge de travail et le temps de récupération favorisant un effet PAP en sprint avec résistance :
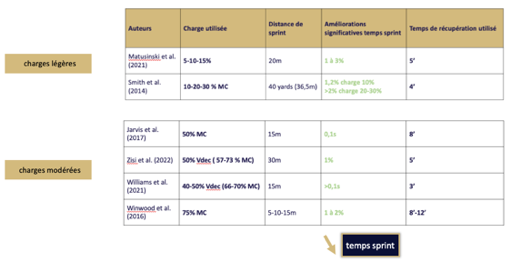
% MC = Pourcentage de masse corporelle
% Vdec = Pourcentage de diminution de la vitesse maximale sans charge
Définition de la PAP :
La Post-Activation-Potentiation (PAP) est définie comme un état neuromusculaire amélioré, observé après l'exécution d'un exercice de haute intensité (Robbins, 2005). C’est un booster neuromusculaire. La PAP se définie par une augmentation de la force de contraction du muscle qui suit une contraction volontaire maximale ou quasi maximale. Le but étant d’entrainer une plus grande production de force si le muscle a été stimulé, activé au préalable.
Les mécanismes physiologiques de la PAP :
D’un point de vue plus physiologique, l’augmentation de la force de contraction musculaire serait liée à une augmentation de la phosphorylation de la chaîne légère de myosine rendant l’actine – myosine plus sensible au calcium CA2+ libéré par le réticulum sarcoplasmique au moment de la contraction musculaire (Hodgson et al., 2005; Sale, 2002). En conséquence, la force de contraction de chaque fibre musculaire s'en voit augmentée. On suppose également que les adaptations de l'activité réflexe dans la moelle épinière, après l'exécution d'un exercice de haute intensité, contribuent à la réponse PAP (Chiu et al., 2003; Guellich & Schmidtbleicher, 1996; Hamada et al., 2000; Scott & Docherty, 2004).
Ainsi, il a été proposé que deux mécanismes physiologiques complémentaires mais intervenant sur différentes fonctions soient responsables de la PAP. L’un est la phosphorylation des chaines légères régulatrice de myosine et l’autre est une augmentation du recrutement des unités motrices d’ordre supérieur. Les deux mécanismes sont complémentaires.
Phosphorylation des chaines légères régulatrice de myosine (RLC) :
Selon l’étude de Tillin & Bishop (2009), une molécule de myosine est un hexamère composé de deux chaînes lourdes (figure 1). Les extrémités aminées de chaque chaîne lourde, classées comme tête de myosine, contiennent deux RLC, et chaque RLC possède un site de liaison spécifique pour l'incorporation d'une molécule de phosphate. La phosphorylation des RLC est catalysée par l'enzyme kinase de la chaîne légère de myosine, qui est activée lorsque les molécules de Ca2+, libérées du réticulum sarcoplasmique pendant la contraction musculaire, se lient à la calmoduline, une protéine régulatrice du calcium.
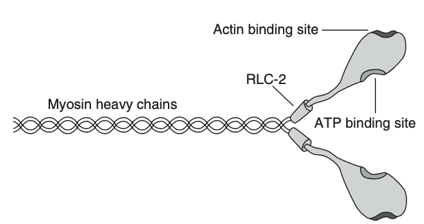
Figure 1 : Une molécule de myosine. Chaque molécule de myosine est composée de deux chaînes lourdes de myosine. La chaîne légère régulatrice (RLC)-2 représente une paire de RLC positionnées au niveau du cou d'une tête de myosine. Chaque RLC peut incorporer une molécule de phosphate, ce qui modifie la structure de la tête de la myosine. Chaque tête de myosine comporte un site de liaison à l'actine et à l'adénosine triphosphate (ATP). (Tillin & Bishop, 2009).
Les chercheurs suggèrent que la phosphorylation du RLC potentialise les contractions subséquentes en modifiant la structure de la tête de la myosine et en l'éloignant de son filament épais. La phosphorylation du RLC a son plus grand effet à des concentrations relativement faibles de Ca2+. Relativement peu d'études ont tenté de mesurer la réponse PAP dans le muscle squelettique humain mais Stuart et al. (1988) ont enregistré une teneur en phosphate significativement élevée du RLC dans le muscle vaste latéral (p <0,01), et une potentialisation significative de la tension de commutation des extenseurs du genou, après une contraction volontaire maximale isométrique de 10 secondes (MVC ; p <0,05). Il y avait également une corrélation positive mais non significative entre l'étendue de la potentialisation de la contraction et la quantité de phosphate incorporée dans les unités RLC individuelles, et entre la potentialisation et le pourcentage de fibres musculaires de type II (p >0,05). (Stuart et al., 1988)
Recrutement des unités motrices d'ordre supérieur :
Les recherches ont montré qu'une contraction isométrique provoquée par la stimulation de fibres neurales afférentes spécifiques, qui activent à leur tour les a motoneurones adjacents via une augmentation neuronale, augmente la transmission des potentiels d'excitation à travers les jonctions synaptiques au niveau de la moelle épinière. Cet état d'accommodation peut durer plusieurs minutes après la contraction isométrique et il en résulte une augmentation des potentiels post-synaptiques pour le même potentiel pré-synaptique lors d'une activité ultérieure. Les chercheurs pensent qu'une contraction musculaire induite diminue la défaillance du transmetteur au cours de l'activité suivante, par l'intermédiaire d'une ou d'une combinaison de plusieurs réponses possibles. Il s'agit notamment d'une augmentation de la quantité de neurotransmetteur libéré, d'une augmentation de l'efficacité du neurotransmetteur ou d'une réduction de la défaillance du point de ramification axonal le long des fibres neuronales afférentes. (Tillin & Bishop, 2009).
D’autres études ont permis d’étudier les effets d’une contraction de conditionnement volontaire (CC) sur le recrutement des motoneurones. Une onde est enregistrée au niveau des fibres musculaires à l'aide de l'électromyographie suite à la contraction présentant une diminution de la défaillance du transmetteur au niveau des jonctions synaptiques et une augmentation subséquente du recrutement des motoneurones d'ordre supérieur.
Guellich & Schmidtbleicher (1996) ont stimulé le nerf tibial et mesuré les changements d'amplitude de l'onde H (réflexe) au niveau du gastrocnémien avant et après cinq contractions volontaires maximales (CMV) isométriques de 5 secondes en flexion plantaire. Ils ont constaté une dépression de l'amplitude de l'onde H 1 minute après les CMV (-24% ; p < 0,05), mais une potentialisation de l'amplitude de l'onde H 5-13 minutes après les CMV (+20% ; p < 0,01).
Modification de la courbe force-vitesse :
Dans la plupart des activités sportives explosives, la puissance joue un rôle majeur et la puissance mécanique peut être définie comme la vitesse à laquelle la force est développée sur une plage de mouvement, dans une période de temps spécifique, ou comme la force multipliée par la vitesse. L'augmentation du niveau de force à une vitesse donnée permet d’augmenter la puissance mécanique. Bien que la PAP semble avoir peu d'effet aux extrêmes de la courbe force-vitesse (figure 2), il a été démontré qu'elle augmentait le taux de développement de la force (RFD) des contractions musculaires. Une augmentation du RFD entraîne une courbe force-vitesse moins concave.
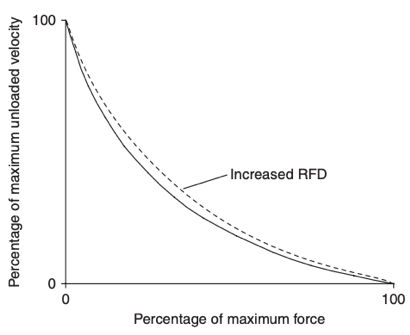
Figure 2 : La relation force vitesse modifiée par la PAP. (Sale, 2002; Tillin & Bishop, 2009)
Ainsi, la PAP pourrait être bénéfique à la performance des activités sportives explosives en augmentant le RFD et donc la puissance mécanique.
L’influence de la période de récupération entre l’AC et la performance suivante sur la PAP :
Le temps de récupération entre l’échauffement PAP et l’activité de puissance suivante est un facteur important qui influence la réponse de potentiation. Il faut choisir un temps suffisamment long mais pas trop pour permettre aux effets de la fatigue de se dissiper sans perdre la potentialisation ou l'effet PAP (Hodgson et al., 2005; Weber et al., 2008).
Si la fatigue est trop importante, par exemple si le sprint avait lieu immédiatement après le mouvement de PAP, alors la PAP ne pourrait pas avoir d'effets optimaux (Robbins, 2005). Et de même, si trop de temps s'écoule, la fatigue diminue, mais les effets de la PAP aussi.
Plusieurs études ont examiné l'impact de différents temps de récupération sur la performance des activités de puissance. Ces études ont trouvé des impacts positifs sur la performance des activités de puissance avec des périodes de récupération allant de 0 à 20 minutes(Bevan et al., 2010; Comyns et al., 2006; Docherty et al., 2004; Kilduff et al., 2007).
Une période de repos appropriée après un échauffement PAP peut également dépendre des activités concernées. Par exemple, Linder et al. (2010) ont constaté une amélioration des temps de sprint sur 100 m après que les participants aient bénéficié d'un temps de récupération de 9 minutes. Des études sur 30m, 40m ont utilisé un temps de récupération de 4 à 5 minutes (Chatzopoulos et al., 2007; McBride et al., 2005). Ces études ont utilisé un squat lourd comme mouvement PAP dans l'échauffement. Bien que plusieurs chercheurs aient examiné le concept d'échauffement PAP, la plupart des études ont porté sur des exercices tels que les squats et les sauts verticaux, plutôt que sur des épreuves de compétition comme le sprint. Les études qui ont démontré un effet significatif de l'échauffement PAP sur la performance du sprint ont utilisé une période de repos de 4 à 10 minutes.
L’équilibre entre la PAP et la fatigue et son effet sur les contractions explosives ultérieures ont été observé et ont rapporté que la fatigue semble plus dominante dans les premiers stades de la récupération et, par conséquent, la performance de l'activité volontaire subséquente est diminuée ou inchangée. Cependant, la fatigue s'estompe plus rapidement que la PAP, et une potentialisation des performances peut être réalisée à un moment donné de la période de récupération. La figure 3 illustre la relation PAP-fatigue.
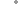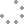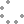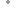
Figure 3 : Modèle de la relation entre la potentialisation post-activation (PAP) et la fatigue à la suite d'un protocole de contraction de pré-conditionnement. Lorsque le volume de la condition est faible, la PAP est plus dominante que la fatigue, et une potentialisation de la performance explosive ultérieure (post/pre) peut être réalisée immédiatement (window 1). Au fur et à mesure que le volume de la condition augmente, la fatigue devient dominante, affectant négativement les performances ultérieures. Après la condition, la fatigue se dissipe plus rapidement que la PAP, et une potentialisation de la performance explosive ultérieure peut être réalisée à un moment donné pendant la période de récupération (window 2). (Tillin & Bishop, 2009).
Si les contractions isométriques et dynamiques peuvent induire des réponses différentes à la fatigue, on peut supposer qu'elles peuvent également avoir des effets différents sur les mécanismes de la PAP : les CMV isométriques peuvent induire une fatigue centrale, mais une PAP périphérique, tandis que les CMV dynamiques peuvent induire la réponse inverse.
Relation entre PAP et fibres musculaires :
Une amélioration des performances grâce à un échauffement PAP dépend aussi de la composition des fibres musculaires. Plus les athlètes ont un pourcentage élevé de fibres musculaires à contraction rapide ou de type II, plus ils présentent des réponses plus importantes à la PAP tandis que les athlètes ayant un pourcentage plus élevé de fibre lente, de type I sont moins sujet à répondre à cet effet PAP. (Hamada et al., 2000). Les fibres II produisent une plus grande quantité de phosphorylation de la chaîne légère de la myosine face à une activité intense, ce qui rend l’actine-myosine plus sensible au calcium lors des contractions musculaires suivantes (Zhi et al., 2005).
Les individus les plus susceptibles de bénéficier de la PAP sont ceux qui ont une plus grande force musculaire, un pourcentage plus élevé de fibres de type 2 (bien que la fatigue puisse également être plus importante chez ces individus), et un rapport puissance-force plus faible, en effet, les sujets les moins capables de convertir efficacement leur force en puissance sont plus susceptibles de bénéficier de la PAP que ceux qui le peuvent.
Notes et références :
Bevan, H., Cunningham, D., Tooley, E., Owen, N., Cook, C., & Kilduff, L. (2010). Influence of Postactivation Potentiation on Sprinting Performance in Professional Rugby Players. Journal of strength and conditioning research / National Strength & Conditioning Association, 24, 701‑705. https://doi.org/10.1519/JSC.0b013e3181c7b68a
Chatzopoulos, D., Michailidis, C., Giannakos, A., Alexiou, K., Patikas, D., Antonopoulos, C., & Kotzamanidis, C. (2007). Postactivation Potentiation Effects After Heavy Resistance Exercise on Running Speed. Journal of strength and conditioning research / National Strength & Conditioning Association, 21, 1278‑1281. https://doi.org/10.1519/R-21276.1
Chiu, L. Z. F., Fry, A. C., Weiss, L. W., Schilling, B. K., Brown, L. E., & Smith, S. L. (2003). Postactivation potentiation response in athletic and recreationally trained individuals. Journal of Strength and Conditioning Research, 17(4), 671‑677. https://doi.org/10.1519/1533-4287(2003)017<0671:ppriaa>2.0.co;2
Comyns, T. M., Harrison, A. J., Hennessy, L. K., & Jensen, R. L. (2006). The optimal complex training rest interval for athletes from anaerobic sports. Journal of Strength and Conditioning Research, 20(3), 471‑476. https://doi.org/10.1519/18445.1
Docherty, D., Robbins, D., & Hodgson, M. (2004). Complex Training Revisited : A Review of its Current Status as a Viable Training Approach. Strength & Conditioning Journal, 26. https://doi.org/10.1519/1533-4295(2004)026<0052:CTRARO>2.0.CO;2
Guellich, A., & Schmidtbleicher, D. (1996). MVC-induced short-term potentiation of explosiv force. New Studies in Athletics, 11, 67‑84.
Hamada, T., Sale, D. G., MacDougall, J. D., & Tarnopolsky, M. A. (2000). Postactivation potentiation, fiber type, and twitch contraction time in human knee extensor muscles. Journal of Applied Physiology, 88(6), 2131‑2137. https://doi.org/10.1152/jappl.2000.88.6.2131
Hodgson, M., Docherty, D., & Robbins, D. (2005). Post-activation potentiation : Underlying physiology and implications for motor performance. Sports Medicine (Auckland, N.Z.), 35(7), 585‑595. https://doi.org/10.2165/00007256-200535070-00004
Kilduff, L., Bevan, H., Kingsley, M., Owen, N., Bennett, M., Bunce, P., Hore, A., Maw, J., & Cunningham, D. (2007). Postactivation Potentiation in Professional Rugby Players : Optimal Recovery. Journal of strength and conditioning research / National Strength & Conditioning Association, 21, 1134‑1138. https://doi.org/10.1519/R-20996.1
Linder, E. E., Prins, J. H., Murata, N. M., Derenne, C., Morgan, C. F., & Solomon, J. R. (2010). Effects of preload 4 repetition maximum on 100-m sprint times in collegiate women. Journal of Strength and Conditioning Research, 24(5), 1184‑1190. https://doi.org/10.1519/JSC.0b013e3181d75806
McBride, J. M., Nimphius, S., & Erickson, T. M. (2005). The acute effects of heavy-load squats and loaded countermovement jumps on sprint performance. Journal of Strength and Conditioning Research, 19(4), 893‑897. https://doi.org/10.1519/R-16304.1
Robbins, D. W. (2005). Postactivation potentiation and its practical applicability : A brief review. Journal of Strength and Conditioning Research, 19(2), 453‑458. https://doi.org/10.1519/R-14653.1
Sale, D. G. (2002). Postactivation potentiation : Role in human performance. Exercise and Sport Sciences Reviews, 30(3), 138‑143. https://doi.org/10.1097/00003677-200207000-00008
Scott, S., & Docherty, D. (2004). Acute Effects of Heavy Preloading on Vertical and Horizontal Jump Performance. Journal of strength and conditioning research / National Strength & Conditioning Association, 18, 201‑205. https://doi.org/10.1519/R-13123.1
Stuart, D. S., Lingley, M. D., Grange, R. W., & Houston, M. E. (1988). Myosin light chain phosphorylation and contractile performance of human skeletal muscle. Canadian Journal of Physiology and Pharmacology, 66(1), 49‑54. https://doi.org/10.1139/y88-009
Tillin, N., & Bishop, D. J. (2009). Factors Modulating Post-Activation Potentiation and its Effect on Performance of Subsequent Explosive Activities. Sports medicine (Auckland, N.Z.), 39, 147‑166. https://doi.org/10.2165/00007256-200939020-00004
Weber, K. R., Brown, L. E., Coburn, J. W., & Zinder, S. M. (2008). Acute effects of heavy-load squats on consecutive squat jump performance. Journal of Strength and Conditioning Research, 22(3), 726‑730. https://doi.org/10.1519/JSC.0b013e3181660899
Annexes :
Tableau récapitulatif de la charge de travail et du temps de récupération favorisant un effet PAP en sprint avec résistance :
% MC = Pourcentage de masse corporelle
% Vdec = Pourcentage de diminution de la vitesse maximale sans charge
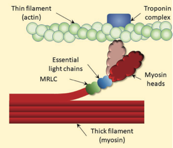
Explication du phénomène de Post Activation Potentiation (Blazevich & Babault, 2019) :
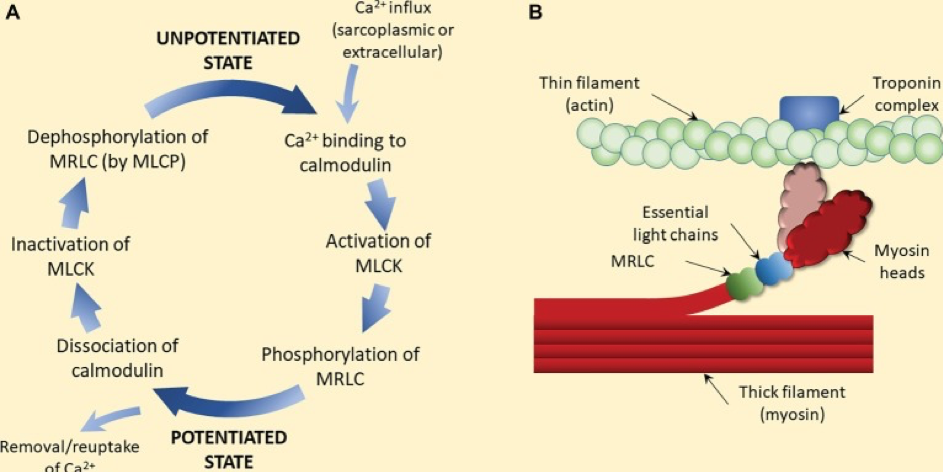
(A) Les ions calcium activent la « myosine light chain kinase » (MLCK) : chaîne légère de myosine suite à l'interaction calcium-calmoduline. La MLCK phosphoryle ensuite la chaîne légère régulatrice de la myosine (MRLC), ce qui est censé déclencher la rotation de la tête de myosine à l'écart du filament épais (squelette de la myosine) vers le filament fin (tête de myosine de couleur claire dans le panneau B). Le retrait ou la réabsorption du calcium déclenche la dissociation de la calmoduline et l'inactivation de la MLCK. La déphosphorylation de la MRLC par la myosine light chain phosphatase (MLCP) complète le processus de suppression de la potentialisation. (B) La phosphorylation de la MRLC et la rotation subséquente de la tête de la myosine (couleur claire) augmentent la probabilité que la tête s'attache à l'actine, et donc la production de force. À des niveaux sous-maximaux (c'est-à-dire inférieurs à la saturation) de calcium, ce processus augmente la production de force à une concentration de calcium donnée, c'est-à-dire la sensibilité au calcium. Comme la phosphorylation est rapide mais que la déphosphorylation est relativement lente, une période d'activité contractile (par exemple, une période d'échauffement) est suffisante pour augmenter les propriétés contractiles lors des contractions suivantes, même si une courte période de repos est imposée.